| • | Biosphere 2 Center (Arizona) |
| • | Black Rock Forest (New York) |
Module 7: Consumers: Herbivory, Predation, and Parasitism |
ConsumersBy Dr. James A. Danoff-Burg, Columbia UniversityAfter learning a bit about producers during Module 6, which are the basis of most ecosystems, we spend today studying the various types of animals that are consumers of live tissue. Consumers consume tissue of either plants or other animals and there are many types. The producer level serves to introduce novel tissue and energy into the ecosystem. Some of the producers are then consumed by a herbivore, metabolized, and used to make new tissue, to move, to respire, and to thermoregulate. This consumptive process continues through all the connections of the community, from herbivores, to primary carnivores (who feed on the herbivores), to secondary carnivores (who feed on the primary carnivores), to tertiary carnivores (who feed on the secondary carnivores), and sometimes to still higher levels of carnivory. Along each of these stages, these consumer levels can all be subject to parasites, parasitoids, mutualists, allelopaths, and commensals. (See Water on the Web's glossary of aquatic ecology terms for definitions of these terms.) We will explore these categories further in a moment. The consumption of dead tissue is completed by the decomposers, a subset of which are also known as detritivores — a group that we will study in depth during Module 8. All community members that consume tissue are referred to as heterotrophs (literally, "other feeders"). In contrast producers are autotrophs ("self feeders") In the past, relationships among species in a community were summarized with a food chain, a linear order of energy transfer between these trophic levels. These linear relationships implied a unidirectional flow of energy and tissue between all the interactors, beginning with the producers, through all the consumer levels, and ending with the decomposers. We have since expanded this concept. The structure of most communities is highly convoluted, with most levels interacting with many individuals across many levels. Therefore, the term food web and its connotations of multiple links between our community members is more appropriate.
Energy Transfer Between ConsumersFor the sake of simplicity, let us return to the more simple and linear food chain. Using this chain, we can make some predictions about how much of the original energy introduced into the community by the producers is eventually available to the terminal consumer (e.g., the tertiary carnivore). At each feeding link in our food chain, we have a loss of anywhere from 70% to 99% of the energy that was available at the lower trophic level, with an average loss of 90%. Why is this? As we said earlier, energy that is released from the consumption and metabolism of tissue is used during movement, respiration, and thermoregulation (in endothermic organisms). These metabolic costs detract from the amount of energy that can be converted into tissue at higher trophic levels. Therefore, about only 1 / 10,000 of the original solar energy captured by the producers would be represented in a food chain ending with a tertiary carnivore. This is part of the reason why some advocate for vegetarianism - much less food is necessary to feed the same number of people.
Types of ConsumersWe discussed the types of consumers that may exist above. This table also lists the benefits or detriments that are inflicted on each species that is involved in the interaction. A plus sign (+) denotes that the species is benefited by the interaction, two plus signs (++) that the interaction is necessary for the existence of that species, minus signs (-) indicate that the species is harmed, two minus (--) signs indicate that the species dies as a consequence, and a zero (0) indicates that the species is not affected by the interaction. 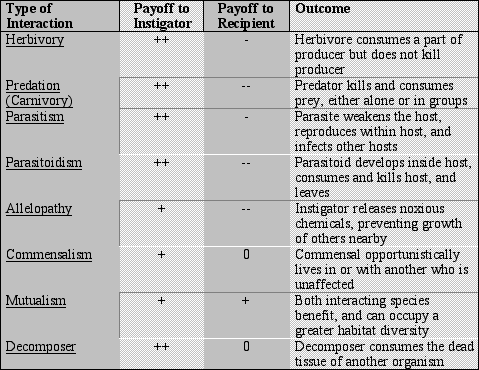
Most of these relationships involve one species benefiting (the user species; e.g., the predator or herbivore) while another is harmed (the supplier species; e.g., the prey or plant) from the interaction. As such, the types of equations used to model the population growth (Module 4) and impact of each species are very similar and are discussed at length in Chapter 10 of Begon, Harper, and Townsend. Although the exact mechanics of them are beyond the scope of our current discussion, they can be tersely summarized. The user species tends to peak in population size one quarter cycle after the recipient species has reached its maximum. The process then repeats through time. Said differently, the instigator species is usually one quarter of a cycle behind the recipient species. 
MutualistsWe will discuss decomposers at greater length during Module 8, but mutualists deserve a brief special attention. Mutualists tend to be ignored by most ecology texts and classes even though they comprise most of the world’s biomass and are arguably responsible for generating most of the Earth’s species. Most grasses, forbs, and trees are mutualists in that they have a symbiotic (literally, "living together") relationship with a fungus that fixes nitrogen. Similarly, coral reefs and plant pollination occur because of intricate mutualistic relationships. Mutualistic relationships are the only type of relationship where both individuals benefit. Usually this benefit takes the form of allowing each species to live in places where they otherwise would not be capable of doing. Mutualisms are often facultative relationships in that the two species can generally live without the other, although both do better when they are with each other. Many mutualisms are obligate relationships, wherein both species need the other to survive and / or propagate. Some extreme examples of these generalities are found within lichens, a symbiotic mutualism wherein a species of algae lives within a fungus. The many species of lichens (an interesting concept in itself — a new species comes about as a consequence of a mutualism) can live one of the most inhospitable of habitats — the surface of bare rock. Neither the algae nor fungus would be able to live in this habitat without the presence of the other. Additionally, there are some species of lichens that are obligate relationships while other lichen species are facultative relationships and the fungus and algal species can live in the wild without each other. In the case of facultative lichens, when they enter into the mutualism, the algae and fungus are both able to take advantage of a much wider diversity of niches than they would have without the mutualism.
Consumers and BiodiversityThe consumer trophic levels comprise most of the species on the planet. Please explore the Described Species activity in Wilson and Perlman for an interactive exploration of their overwhelming numerical dominance. How did this happen? What is the effect on community diversity if some of the consumer species are removed? How often are keystone species (from Module 5) consumer species?
How can we explain the biodiversity of consumer species? The Long-Legged Ant Beetle (Sceptobius schmitti, typo in image to right) However, environmental disruptions are not essential for the creation of so many consumer species. In the exact same relationship above, we could get the production of additional species merely due to increasing specialization. Let’s say that initially the beetle did not specialize on that species of ant and lived in the leaf litter, occasionally venturing into the nests of many ant species and taking food and temporary shelter from them all. Through time, several of the beetle lineages began to prefer to live with a different species of ant. As a result they became better able to exploit the host ant. In return, the ant began to fight them off and try to keep them out of the nest, which the beetle quickly countered. These back-and-forth changes in response to each other continued and resulted in permanent alterations in the behavior, morphology, and genetic composition of both the ant and beetle species. As a consequence, the interacting species became specialized only on each other and many additional species of beetles and ants were produced in other parts of the range of these insects. This process of back-and-forth change and increasing specialization, resulting in speciation is a process called coevolution. Coevolution plays a large role in generating consumer species. Consumer species are often specialists in that they feed on (or live with, in the case of mutualists) one or very few species. Specialization frequently leads to speciation, which is how many consumer species may have been produced. As we mentioned during Module 6, many species of plants engage in a coevolutionary chemical arms race with their insect herbivores. Much of the evolution of parasite and parasitoid species has come about as a consequence of their hosts speciating. The list of coevolutionary relationships is tremendously large. However, many species feed more generally and are therefore oligotrophs. Oligotrophs will not feed on everything, as will a generalist, but it is still less clear how they could produce a great number of species. Coevolution is less likely when the relationships between the species are more diffuse as in the case of oligotrophs. Predators are typically oligotrophic and feed on many species of animals. It is possible that predators are not very species rich because of this, in relation to other consumer species. Consumers help to increase local biodiversity on a much-shorter ecological time scale as well. Some work exploring the impact of a predaceous starfish off the Washington State coast found that when the starfish was removed from the ecosystem, the number of species present halved and the number of trophic levels reduced. Interestingly, the overall standing crop (Module 6) increased when the starfish was removed. These facts were explained because when the terminal predator (the starfish) was removed, it allowed one barnacle species to explode in population size and exclude other species. When the starfish was present it fed on the most common species (a phenomenon called prey switching), kept all of the species in check, and thus allowed a greater diversity of prey species to coexist. Therefore, consumers comprise the majority of the extant species and interrelationships and through their activities they further enhance local biodiversity. There are so many species of consumers that the types of questions that could be answered with their assistance are nearly inconceivable.
Additional Relevant Online ResourcesOrnithology.com is both a central clearinghouse for information about birds, including a Water on the Web from the University of Minnesota has an excellent glossary of aquatic ecology terms. The Soil Biology Primer, from the USDA's NRCS Soil Quality Institute in Ames, Iowa, has many chapters on soil ecology. Dr. Michael J. Pidwirny, of the Department of Geography, Okanagan University College, has an excellent online textbook on Physical Geography. The Introduction to Biogeography and Ecology chapter is particularly relevant to communities. Davey Jones of the University of Wales, Bangor, has a thorough lecture page on Lichens. Fun with Lichens from Oregon State University has information on the basic biology of these fascinating organisms, as well as an interactive identification key to lichens.
All Materials Copyright © 2000 by James Danoff-BurgAll Rights Reserved.
|
 lives with and takes food and shelter from the Velvety Tree Ants (Liometopum luctuosum) in southeastern Arizona. These symbiotic beetles have never been collected from anywhere else aside from inside the nest of that ant species. As a consequence, were the ants to speciate into two species, say in response to an environmental change that disrupts their otherwise continuous distribution range, then the beetles would eventually speciate as well. In this evolutionary scenario involving an obligate specialist relationship, it is clear how consumers could have become so species-rich. When the host species speciates, the guest does as well.
lives with and takes food and shelter from the Velvety Tree Ants (Liometopum luctuosum) in southeastern Arizona. These symbiotic beetles have never been collected from anywhere else aside from inside the nest of that ant species. As a consequence, were the ants to speciate into two species, say in response to an environmental change that disrupts their otherwise continuous distribution range, then the beetles would eventually speciate as well. In this evolutionary scenario involving an obligate specialist relationship, it is clear how consumers could have become so species-rich. When the host species speciates, the guest does as well.